

Le piante hanno tre modi di difendersi dalle malattie parassitarie
- evitandole: ad es. portando a maturazione i frutti prima dell’esplosione epidemica.
- tollerandole: la tolleranza è da intendersi come la capacità di sopportare la malattia , valutabile in base alla stessa produzione, o come capacità di sopportare il patogeno senza manifestazioni patologiche. Abbiamo a questo proposito TOLLERANZA DELLA MALATTIA con meccanismi di compensazione da parte delle parti non colpite dalla malattia, e TOLLERANZA DEL PARASSITA quando la presenza del parassita non determina malattia.
- contrastandole: la resistenza vera e propria si manifesta con la capacità di contrastare il parassita. Anche in questo caso ci troviamo di fronte a RESISTENZA GENERALE che tutte le piante esercitano verso saprofiti e patogeni eterologhi, RESISTENZA SPECIFICA O VARIETALE esercitata contro patogeni omologhi (specifici), come lo spessore della
4. cuticola, secrezione di composti antimicrobici. I meccanismi di resistenza si distinguono in passivi preinfezionali e precostitutivi, e attivi postinfezionali e indotti. Una ulteriore distinzione viene fatta:
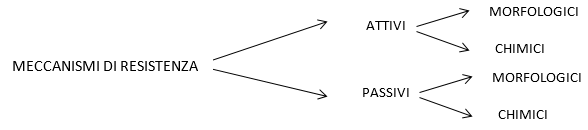
MECCANISMI DI RESISTENZA PASSIVI
Morfologici passivi:
Contrasto dell’adesione e dello sviluppo
- organotropia sfavorevole al crearsi di umidità elevata
- cuticole e pelosità idrorepellenti
- microscanalature cunicolari che disturbano l’orientamento verso gli stomi e la penetrazione dei tubuli germinativi
Contrasto della penetrazione e successivi processi di colonizzazione
- presenza di sughero
- spessore e lignificazione delle pareti cellulari
- lenticelle piccole o poco numerose
- stomi non differenziati
- scarsa pervietà degli organi fiorali e dei frutti
Chimici passivi:
- assenza sulle superfici di composti che attraggono i patogeni
- assenza di composti formativi
- assenza di principi nutrizionali indispensabili per la crescita e sviluppo dei patogeni
- condizioni inadatte di ph
- concentrazioni di zucchero troppo elevate
- presenza di composti in grado di neutralizzare enzimi e tossine dei patogeni
- presenza di composti ad azione antimicrobica indiretta (proibitine)
- presenza nelle pareti di composti in grado di catturare i patogeni (lectine) In virtù di queste difese le piante possono acquisire proprietà refrattarie verso i patogeni e molti organismi estranei
MECCANISMI DI DIFESA ATTIVI
Comportano reazioni metaboliche molto complesse a carico di cellule , tessuti e a volte anche dell’intera pianta.
Morfologici attivi:
- risposte di difesa cellulare ( citoplasmatico ), come ispessimenti di parete direttamente sollecitata con depositi di calloso beta1,3-glucano, oppure apposizione di strati attorno all’ifa infettiva avanzante con formazione di forme coniche rivolte verso il lume cellulare chiamate papille o lignituberi. Produzione di gomme in quantità molto variabile da specie a specie impregnate di tannini (antibiotici) negli spazi intercellulari e nelle stesse cellule. Produzione di gomme o di tille nei vasi legnosi da parte di cellule parenchimatiche annesse ai vasi rappresentano una barriera molto efficace alla diffusione nello xilema di parassiti vascolari. Difese più complesse sono rappresentate da formazioni simili a cicatrizzazione con differenziazione di fellogeno attorno a punti invasi difficilmente superabili dal patogeno perché impermeabile alle tossine ed agli enzimi prodotti dal parassita; spesso a queste formazioni segue il distacco della parte colpita favorendo l’eliminazione fisica del patogeno.
Chimici attivi:
- sintesi e accumulo di composti di natura aromatica (antimicrobici) postinfezionale; ciò spinge la pianta ad una maggiore attivazione della respirazione (ciclo respiratorio dei pentosi-fosfati). Grande importanza come fattori di difesa viene attribuita alle fitoalessine, dotati di forte attività antimicrobica , che si formano in seguito ad inoculazione o contatto con microrganismi. Le fitoalessine vengono prodotte soprattutto da piante resistenti più che in quelle suscettibili. Mentre le fitoalessine vengono prodotte localmente, la neosintesi di enzimi capaci di lisare la parete di funghi e batteri viene indotta sistematicamente in organi distanti da quelli inoculati o danneggiati.
REAZIONE DI IPERSENSIBILITA’
Consiste in una rapida risposta dell’ospite all’infezione in genere risultante dalla morte del tessuto invaso con obiettivo preventivo alla diffusione del patogeno. Dipende da caratteri di estrema sensibilità dell’organismo agli stimoli del patogeno. La morte di poche cellule , seppur trascurabile per l’intera pianta, diventa funzionale alla sopravvivenza della pianta stessa. L’imbrunimento e la necrosi in genere seguono alterazioni biochimiche e morfologiche della pianta (deposito di lignina, sughero, accumulo di fitoalessine) che si verificano nelle aree immediatamente circostanti. In questo modo i parassiti vengono uccisi o almeno bloccati.
IL RICONOSCIMENTO
I fisiopatologi vegetali sono prioritariamente impegnati a chiarire le modalità con le quali la pianta riesce a percepire “l’invasore” come estraneo (not self), e quindi trattato come incompatibile e come invece percepisca come proprio un tessuto o un microrganismo compatibile. L’evento di riconoscimento si verifica sin dalle prime fasi del processo infettivo ed è facilitato dalla presenza degli elicitori (che provocano) che innescano nella pianta meccanismi di difesa e resistenza ed in particolare accumulo di fitoalessine, reazioni di ipersensibilità, lignificazione, formazione di papille, ecc.
ELICITORI GENERALI, ESOGENI ED ENDOGENI
- I lucani sono elicitori rilasciati dalle pareti di funghi del genere phytophtora e
collectotricum - Chitina e chitosano sono i componenti principali della parete di ascomiceti e basidiomiceti e sono risultati elicitori di fitoalessine in pisello, grano e riso. Le chitinasi attivate dall’infezione agiscono sulle pareti cellulari di Fusarium solani.
- Acidi grassi insaturi di cui sono ricche le pareti di phytophtora sono in grado di indurre produzione e accumulo di fitoalessine nella patata, e di indurre resistenza in piante di pomodoro alle tracheofusariosi.
- Proteine e glicoproteine sono stati individuati elicitori di questo tipo in puccinia e phytophtora ( in varie specie di phytophtora fungono sia da elicitori che da tossine).
- Oligogalatturonidi fungono da messaggeri secondari con un’induzione a distanza.
ELICITORI SPECIFICI
L’adozione di tecniche di biologia molecolare e ingegneria genetica ha consentito una maggiore comprensione dei meccanismi di riconoscimento e della resistenza varietale regolata dal rapporto gene-a-gene.
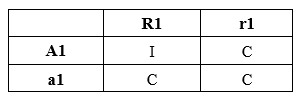
L’analisi teorica della relazione gene-a-gene mediante il confronto in quadrato (quadratic check) portava a concludere che tre diverse combinazioni genotipiche delle quattro possibili risultano con fenotipo suscettibile e una sola (A1/R1) presenta fenotipo resistente. Ciò indica che la resistenza è l’incompatibilità prodotta dall’interazione tra il prodotto del gene A1(composto elicitore) e il prodotto del gene R1(composto ricettore) con la formazione di un prodotto che inibisce il patogeno o tali da pilotare una serie di eventi che culminano nella resistenza (RI). La ricerca di elicitori specifici e dei relativi geni:
- Nel TMV-tabacco l’induttore è stato riconosciuto nel capside proteico
- In Cladosporium fulvum(agente della cladosporiosi del pomodoro) un elicitore razzaspecifico è stato purificato ed ha dimostrato che la inattivazione del gene dell’avirulenza lo rende virulento anche per cultivar di pomodoro che posseggono il gene di resistenza Cf9.



« Come avviene l’inoculo Da bruco a farfalla: scoprite il miracolo della metamorfosi »

 0422 794002
0422 794002






{kind=link}